Plasmids IS Elements Transposons And Retroelements Plasmids
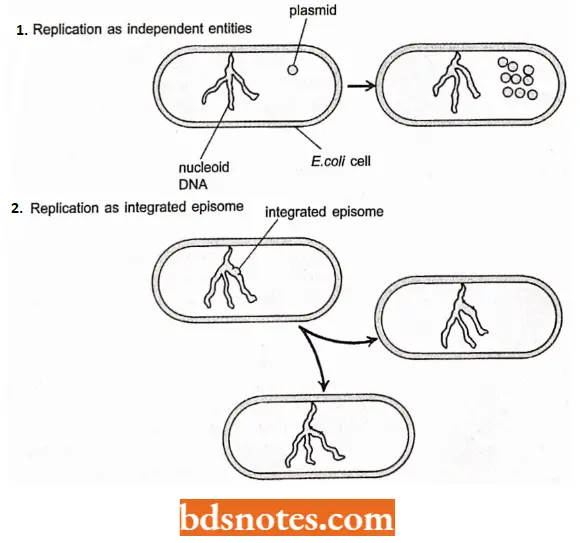
Plasmids are autonomous self-replicating double-stranded circular DNA molecules that are maintained in the cell in a stable and characteristic number of copies.
- A plasmid is a replicon that is stably inherited in an extrachromosomal state. Most, but not all, plasmids are dispensable as they are not essential for the survival of bacterial cells, except under certain environmental conditions.
- The word plasmid was originally used by J. Lederberg (1952) for any extrachromosomal hereditary determinant.
- This term is now restricted to those accessory DNA circles, which are found in bacteria (or in cell organelles such as mitochondria in eukaryotes), in addition to the main chromosome. The nucleoid DNA molecule is not the only repository for genes in the bacterial cell.
- Bacteria often also harbor small independent DNA molecules, the plasmids, which carry genes not found on the main DNA molecule. Plasmids are circular molecules that lead an independent existence in the bacterial cell.
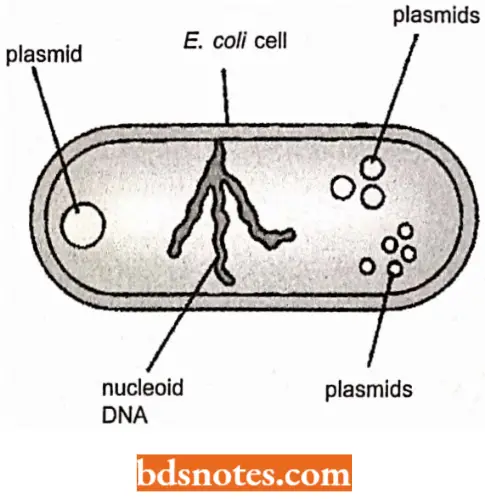
- Plasmids carry one or more genes which are often responsible for a useful characteristic. For example, the ability of bacteria to survive in normally toxic concentrations of antibiotics such as chloramphenicol or ampicillin is often due to the presence of a bacterium of a plasmid that carries antibiotic-resistance genes.
- Genes carried by plasmids also provide bacteria the ability to utilize certain complex compounds such as toluene as a carbon source. All plasmids possess at least one DNA sequence that can act as an origin of replication (replicon), so they can multiply in the cell independently of the main DNA molecule.
- The smaller plasmids make use of the cell’s own DNA replicative enzymes to make copies of themselves, whereas some of the larger ones carry genes that code for special enzymes that are specific for plasmid replication.
- A few types of plasmids are also able to replicate by inserting them into the main DNA molecule. These integrative plasmids are called episomes and they may be stably maintained in this form through numerous cell divisions, but always at some stage exist as independent elements.
Characteristics Of Plasmids
Plasmids share the following general properties:
- They are genetic elements made up of DNA;
- They are smaller than and separate from the main chromosome, and
- They are capable of replication. Some plasmids such as sex factors have unique properties such as
- They control bacterial conjugation,
- They are reversibly inserted in the bacterial chromosome and
- They are transferred from one bacterium to another.
“Understanding plasmids, IS elements, and transposons through FAQs: Q&A explained”
Further, bacteriophages that are included in the category of episomes are sometimes excluded from the category of plasmids, since they show unrestricted replication, leading to lysis. But some phages such as λ (lambda) phages share most of the properties of plasmids and are grouped with them by some workers.
Classification Of Plasmids
Almost all species of bacteria harbor plasmids and a large number of different types are known. They are classified according to the genes they carry and the characteristics that these genes confer on the host bacterium. According to this classification, there are the following five main types of plasmids:
Types Of Special Features Of Some Bacterial Plasmids:

- Fertility(F) plasmids. These plasmids direct conjugation between different bacteria. They are also called conjugative or transmissible plasmids.
- An example is the F plasmid of E. coli. During conjugation of E. coli, when the F+ strain (carrying F factor) comes in contact with the F– strain, two processes may follow:
- One involves the transfer of the F factor so that F– becomes F+ and the second involves the transfer of a segment of the bacterial chromosome, which is facilitated by the insertion of the F factor within the bacterial chromosome at one or other sites, leading to the production of Hfr (high-frequency recombinant).
- Some F plasmids behave like the episomes, i.e., they can exist in a ‘free’ or ‘integrated’ state. Resistance (R) plasmids.
- These plasmids carry genes imparting to the host bacterium the resistance to one or more antibacterial agents such as streptomycin, tetracycline, chloramphenicol, ampicillin, sulphonamide, or mercury.
- R plasmids are very important in clinical microbiology because their spread through natural populations can have acute consequences for the treatment of bacterial infections.
- An example is RP4, commonly found in Pseudomonas but also occurring in other bacteria.
- R plasmids were discovered in Japan in 1956 when a strain of bacteria called Shigella was shown to acquire resistance against several drugs in one step.
- This was shown to be due to the transmission of resistance from one strain to another and was later attributed to R plasmid.
- R plasmid not only confers resistance but also enables bacteria carrying them to conjugate (even in the absence of F factor) and transfer the R plasmid to other bacteria, related or quite unrelated.
- For example, R plasmids of E. coli are known to be transferred to Proteus, Salmonella, Haemophilus, Pasteurella, Shigella, etc.
- Due to further investigations, the genes for drug resistance were found to be a transposon (called Tn3) with terminal inverted repeat (IR) sequences.
Compound R Plasmids: IS sequences and transposable (Tn) elements are responsible for the transfer of genes controlling resistance to antibiotics and other drugs from one genetic element to the other. Such transpositions generate R plasmids having resistance genes for a variety of antibiotics.
Such R plasmids are called compound R plasmids and many such compound R plasmids have been isolated and characterized.
“How do plasmids and transposons contribute to genetic diversity? FAQ answered”
- Sometimes the R plasmids are found to contain two distinct parts: one being genes for the resistance (called R factor) and the other responsible for the transfer of plasmid (called resistance transfer factor or RTF).
- But in some other cases such as penicillin resistance in Staphylococcus, the resistance may not be associated with the ability to transfer (RTF). In such a case transfer of drug resistance may be brought about by transduction.
- Col plasmids. These plasmids carry genes coding for colicins, the toxic proteins that kill other bacteria. An example is the plasmid Col El of E. coli.
- Colicin is produced by Escherichia coli, Shigella, and Salmonella due to their Col plasmids. Col plasmids resemble the F factor or R factor.
- Like the transfer of the R factor, the transfer of the Col factor does not depend on the F factor.
- Since the transfer of the Col factor can occur between two F– strains of bacteria, one carrying Col and the other lacking it. There are, however, some col plasmids (for example., Col El) that require the help of sex factor for its transfer.
- Due to this property, R plasmids and Col plasmids are also called nonconjugative or nontransmissible plasmids.
- Degradative plasmids. These plasmids allow the host bacterium to metabolize unusual molecules such as toluene and salicylic acid. Several examples occur in the Pseudomonas genus of bacteria, for instance, TOL of Pseudomonas putida.
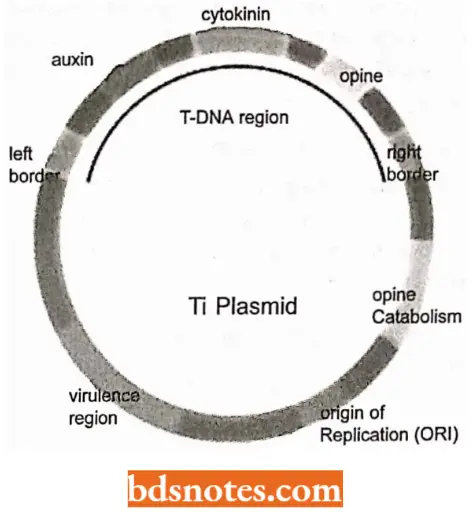
- Virulence plasmids. These plasmids confer pathogenicity on the host bacterium. The best-known example is the Ti plasmid of Agrobacterium tumefaciens, which induces crown gall disease in dicotyledonous plants.
Replication, Transfer, And Recombination In Plasmids: All plasmids discovered so far are small circular DNA molecules. The DNA circles may be single or multiple structures that are linked together in chains.
- The size of these circular DNA molecules varies from 30pm (for example., F+) in the plasmid of E. coli, making 20% of the main chromosome, or can be less than 1 pm as in the P16C of E. coli.
- Colicin plasmids such as Col El, E2, and E3 usually have a length of 2.3 μm. The number of plasmids in a bacterial cell varies from a few in the case of F and R plasmids to as many as 20 – 30 in the case of Staphylococcus plasmids.
Properties Of Various Plasmids:

Copy Number Of Plasmids: Each plasmid is maintained in the bacterial cell at a characteristic copy number mainly due to its replication control system. There are following methods for the regulation of the plasmid’s copy number in a cell:
- Single Copy plasmids. They are maintained in one copy per bacterial cell. Their replication control is the same as that of the bacterial host chromosome so that they replicate and segregate with the bacterial chromosome.
- Such a replication control is called stringent control. Multicopy plasmids. They exist in several, but a characteristic number of copies (typically 10 to 20 copies) per cell.
- Their replication control permits more than one replication to occur while the host cell chromosome replicates once. This type of replication control is called relaxed control; it allows a pool of origins to exist in each cell.
- The DNA of plasmids replicates in a semi-conservative manner. While the initiation of replication is controlled by plasmid genes, other stages of replication such as elongation (polymerization) and termination are controlled by bacterial genes.
- Further, when a plasmid is inserted in the bacterial chromosome, bacterial control on plasmid replication is absolute, but when it is free, the control is less than absolute.
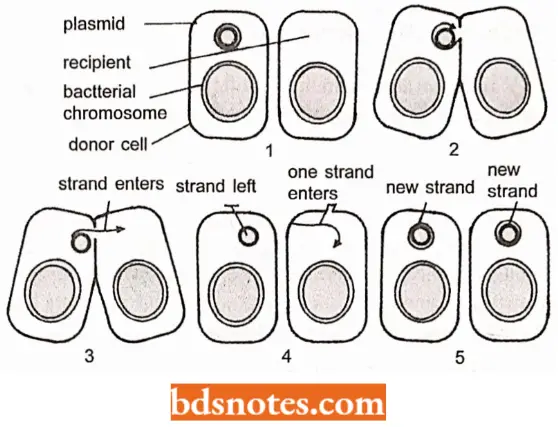
- The property of transfer of plasmid from one bacterium to another is facilitated by the formation of a hair-like extracellular growth, called sex pilus. Sex pilus is formed of the outer membrane of the donor bacterium.
- The F sex pilus is up to 20 μm in length and 8 nm wide. In E. coli, transfer is controlled by plasmid genes tra-A, tra-B, tra-C, etc., and mutations in these genes lead to failure of transfer.
- But, when two plasmids containing mutations for different tra- genes are introduced in the same cell, pilus may be formed and transfer may occur due to complementation between mutants.
- In the process of transfer, one of the two strands of plasmid DNA is nicked (broken) at a site called the origin of transfer (Ori-T) and the linear strand thus formed moves to the recipient bacteria.
- The single strands in the donor as well as in the recipient cell then synthesize complementary strands, so that duplex DNA plasmids are reconstituted in both cells.
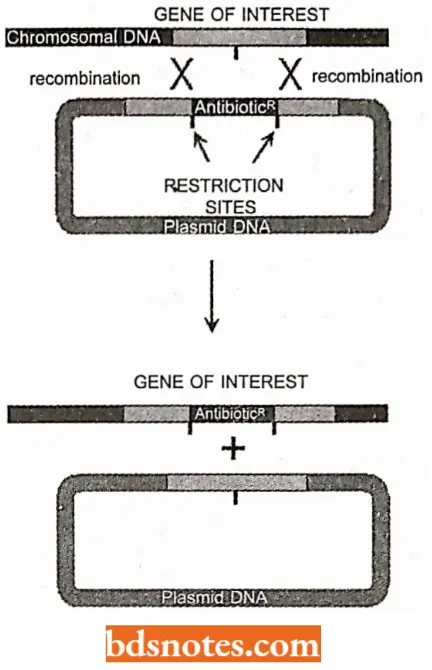
- Lastly, plasmid DNA may undergo recombination either with another plasmid DNA or with bacterial DNA. In the bacterium chromosome, there are insertion sequences (IS), which are homologous in recombination. There are at least 17 such sites m.E. coli genome.
- Mapping of genes in plasmid DNA has also been achieved. Methods employed in the plasmid mapping involved recombination and complementation between strains that have overlapping deletions.
Is Elements And Transposable Elements
A great part of classical genetic analysis has been devoted to the localization of genes on chromosomes. Accordingly, if we map a gene today, we expect to see it in the same place tomorrow.
- However, our discovery of mobile genetic elements has modified that view to some extent.
- We now know that some segments of the genome can move quite readily from one place to another.
- The movement of those elements has effects on the phenotype of the organism, primarily at the transcriptional level.
“Importance of studying plasmids and transposons for biology students: Questions explained”
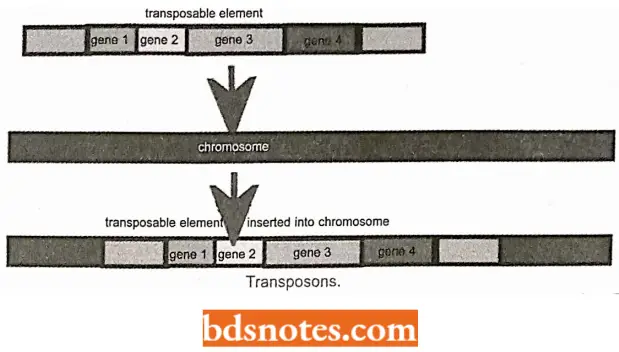
- A transposable element, mobile gene, jumping gene, or transposon is a segment of DNA that can move from one position in a genome to another.
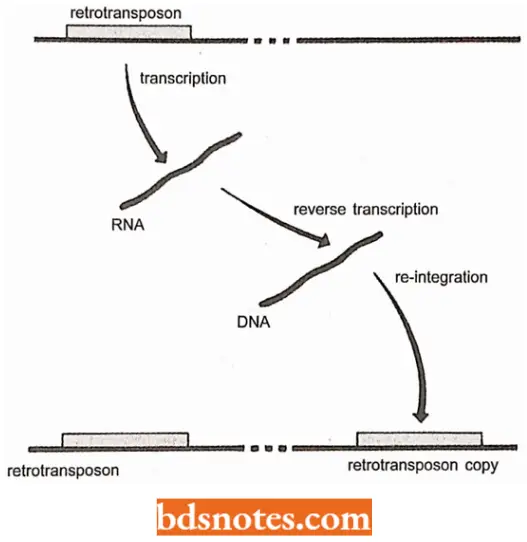
- Retrotransposon is a type of transposable element that moves by being first transcribed into RNA copy that is then reconverted to DNA by reverse transcriptase and inserted elsewhere in the chromosome.
- Thus, a transposable element is a DNA segment that translocates to other sites in the genome, essentially independent of sequence homology.
- Usually, such elements are flanked by short, inverted repeats of 20 to 40 base pairs at each end. Insertion repeats into a structural gene can produce a mutant phenotype.
- Insertion and excision of transposable elements depend on two enzymes, transposase and resolvase. Such transposable elements have been identified in both prokaryotes and eukaryotes.
Discovery And History: In the 1940s, in a study of the genetics of kernel mottling in maize, Barbara McClintock discovered an element that not only regulated the mottling but also caused breakage of the chromosome carrying the genes for color and consistency of the kernel.
- The element was called dissociation (Ds). Mapping data showed that chromosome breakage always occurs at or very near the location of Ds. McClintock’s critical observation was that Ds does not have a constant location but occasionally moves to a new position (transposition).
- Causing chromosome breakage at the new site. Furthermore, Ds moves only if a second element, called activator (Ac) is also present.
- Ac itself moves within the genome and can cause the expression of genes at or near its insertion site, alternations similar to the modifications that result from the presence of Ds. McClintock (1956) referred to Ac and Ds as controlling elements. Both of these form a two-factor system of transposable elements of maize.
- The term transposable elements was coined by another great geneticist Alexander Brink.
- In the mid-1960s Susum Mitsuhashi first suggested that the gene responsible for resistance to several antibiotics was mobile and could move between bacterial plasmids and chromosomes. The first observation of jumping genes at the molecular level came in the early 1970s.
- Several independent researchers, including Peter Starlinger and James Shapiro, visualized a unique class of mutations affecting different genes in various bacterial strains.
- The term transposon was coined by R.W. Hedges and A.E. Jacob in 1974 for a DNA segment that could move from one DNA molecule (or chromosome) to another and carried resistance to the antibiotic ampicillin.
- In 1975, David Hogness and his colleagues David Finnegan, Gerald Rubin, and Michael Young identified a class of mobile genes in Drosophila melanogaster that they designated as Copia.
Transposition And Site-Specific Recombination: In the general type of recombination, DNA rearrangements occur between DNA segments that are very similar in sequence.
- Although these rearrangements can result in the exchange of alleles between chromosomes, the order of the genes on the interacting chromosome typically remains the same.
- A second type of recombination, called site-specific recombination, moves specialized nucleotide sequences, the mobile genetic elements, between nonhomologous sites within a genome.
- Due to this reason, this type of recombination is also called illegitimate recombination (Brown, 1998). Here, movement can occur between two different positions in a single chromosome, as well as between two different chromosomes.
- Unlike general recombination, site-specific recombination is guided by recombination enzymes that recognize short, specific nucleotide sequences present on one or both ends of the recombining DNA molecules.
- Each type of mobile element generally encodes the enzyme that mediates its movement and contains special sites upon which the enzyme acts.
- Many elements also carry other genes. For example, viruses encode coat proteins that enable them to exist outside cells, as well as essential viral enzymes.
- The spread of mobile elements that carry antibiotic-resistance genes is a major factor underlying the widespread distribution of antibiotic resistance in bacterial populations (for example., the spread of resistance for antibiotics such as ampicillin and tetracycline).
- Site-specific recombination mainly occurs by transposition or transpositional site-specific recombination which involves breakage reactions at the ends of the mobile DNA segments embedded in chromosomes and the attachments of those ends at one of many different nonhomologous target DNA sites.
- It does not involve the formation of heteroduplex DNA.
- In transposition, a specific enzyme, usually encoded by the transposon and called a transposase, acts on a specific DNA sequence at each end of the transport — first disconnecting it from the flanking DNA and then inserting it into a new target DNA site.
- There is no need for homology between the ends of the element and the insertion site. Most transposons move only very rarely (once in 10s cell generations for many elements in bacteria).
“Common challenges in understanding plasmids, IS elements, and transposons effectively: FAQs provided”
Types Of Transposons
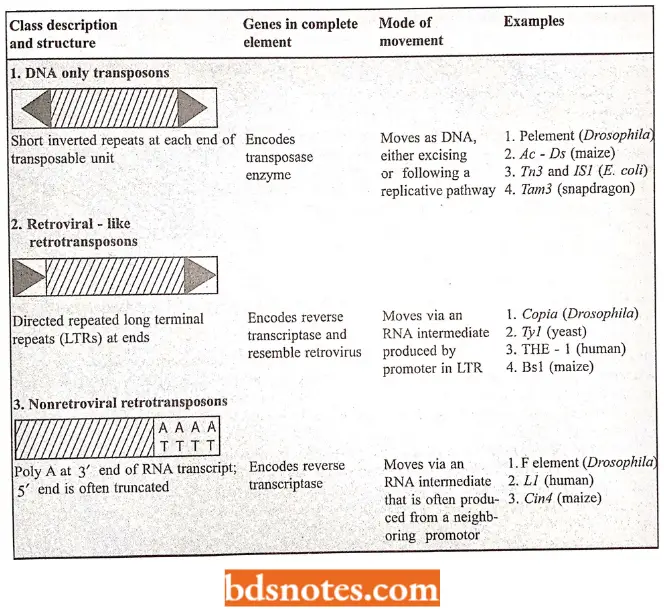
Based on their structure and transposition mechanisms, transposons can be grouped into the following three large classes:
- DNA only transposon. The mobile element exists as DNA throughout its life cycle — the translocating DNA segment is directly cut out of the donor DNA and joined to the target site by a transposase enzyme.
- Retroviral – like retrotransposons. They move by a less direct mechanism. An RNA polymerase enzyme first transcribes the DNA sequence of the mobile element into RNA.
- The enzyme reverse transcriptase then transcribes this RNA molecule back into DNA using the RNA as a template, and it is this DNA copy that is finally inserted into a new site in the genome.
- For historical reasons, the transposase-like enzyme that catalyzes this insertion reaction is called an integrase rather than a transposase.
- Non retroviral retrotransposons. Each of these transposons moves by making a DNA copy of an RNA molecule that is transcribed from it.
- However, the mechanisms involved in these nonretroviral retrotransposons are distinct from those in which the RNA molecule is directly involved in the transposition reaction.
- All three types of transposable elements range in length from 1000 to about 12000 nucleotide pairs; each family contains many members, In addition to trnnsposable elements, there are selected viruses that can move in and out of host cell chromosomes; these viruses are related to the first two classes of transposons.
“Steps to explain types of plasmids: F plasmids vs R plasmids: Q&A guide”
Mode of Transposition of DNA Transposons: DNA transposons may have the following two methods of transposition:
- Replicative transposition. This transposition mechanism involves direct interaction between the donor transposon and the target site resulting in the copying of the donor element, for example., Shapiro’s model.
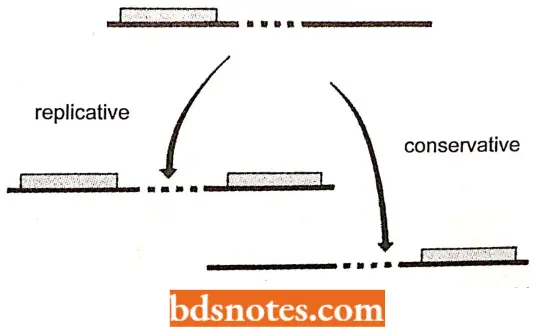
- Conservative transposition. This transposition mechanism involves excision of (lie element and re-intention at a new site, (for example., cut-and-paste model.
- Both mechanisms require enzymes which are usually coded by genes within the transposons. DNA transposons use either the replicative or conservative pathway (some can use both).
“Role of IS elements in genome rearrangement: Questions answered
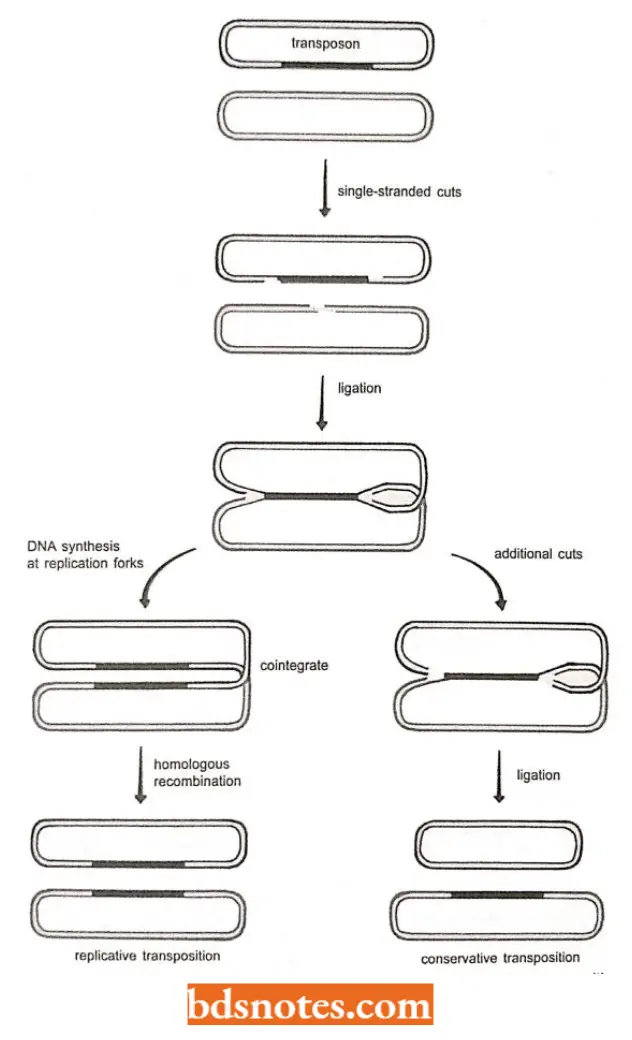
Shapiro’s model of transposition: Several models for replicative and conservative transposition have been proposed over the years but most are modifications of a scheme originally outlined by J.A. Shapiro (1979).
- According to Shapiro’s model, the replicative transposition of a bacterial element such as Tn3- a type transposon or a transposable phage is initiated by one or more endonucleases that make single-stranded cuts on either side of the transposon and in the target site where the new copy of the element will be inserted.
- At the target site, the two cuts are separated by a few base pairs, so that the cleaved double-stranded molecule has short 5′ overhangs.
- Ligation of these 5′ overhangs to the free 3′ ends on either side of the transposon produces a hybrid molecule in which the original two DNAs — the one containing the transposon and the one containing the target site – are linked together by the transposable element flanked by a pair of structures resembling replication forks.
- This step is catalyzed by a transposon-coded transposase enzyme. Repair – DNA synthesis at these replication forks copies the transposable element and converts the initial hybrid into a co-integrate, which contains two transposons and two linked DNAs.
- Now resolvase mediated homologous recombination between the two copies of the transposon uncouples the cointegrate, separating the original DNA molecule (with its copy of the transposon still in place) from the target molecule.
- This process is called resolution. At this stage, the target molecule also contains a copy of the transposon.
- Replicative transposition has therefore occurred. Resolution means the separation of a pair of recombining double-stranded DNA molecules.
- A modification of the process of replicative transposition makes it the conservative transposition. In this case, rather than carrying out DNA synthesis, the hybrid structure is converted back into two separate DNA molecules simply by making additional single-stranded nicks at either side of the transposon.
- This cuts the transposon out of its original molecule, leaving it “pasted” into the target DNA. Thus, conservative transposition involves a “cut-and-paste” mechanism. 2. Cut-and-paste transposition.
- Many DNA-only transposons move from a donor site to a target site by cut-and-paste termination. DNA-only transposons can be recognized in chromosomes by the “inverted repeat DNA sequences” at their ends.
- Experiments show that these sequences, which can be as short as 20 nucleotides, are all that is necessary for the DNA between them to be transposed by the particular transposase enzyme (a dimeric enzyme) associated with the element.
- The cut-and-paste movement of a DNA-only transposable element from one chromosomal site to another brings the two inverted DNA sequences together, forming a DNA loop.
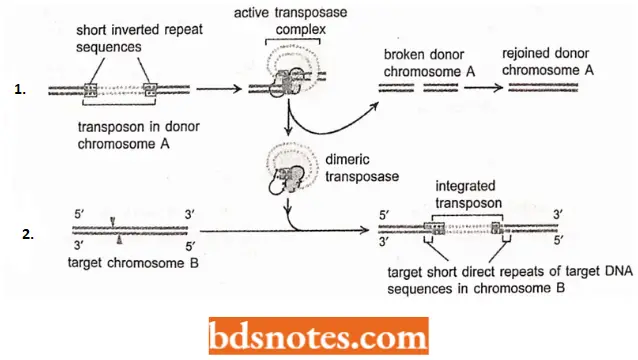
- Insertion into the target chromosome, catalyzed by the transposase enzyme, occurs at random sites through the creation of staggered breaks in the target chromosome.
- As a result, the insertion site is marked by a short direct repeat of the target DNA sequence.
- Since, the breaks made in two target DNA strands are staggered two short, single-stranded gaps are initially formed in the product DNA molecule, one at each end of the inserted transposon.
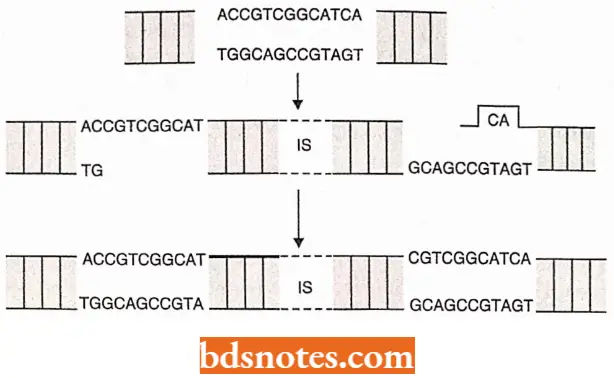
- These gaps are filled in by a host cell DNA polymerase and DNA ligase to complete the recombination process, producing a short duplication of the adjacent target DNA sequence.
- These flanking direct repeat sequences serve as convenient markers of a prior transpositional site-specific recombination event.
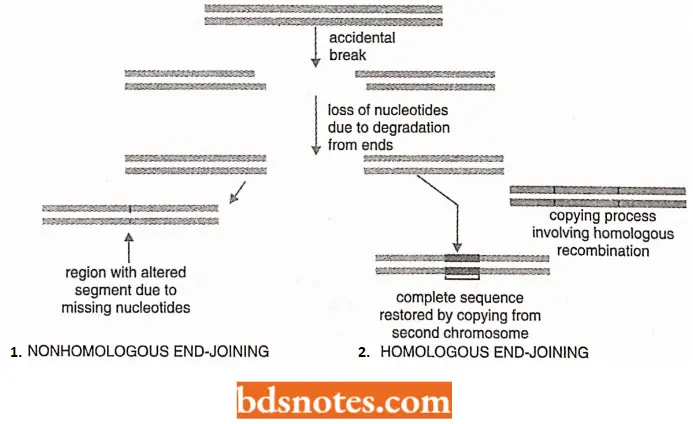
- Further, when a cut-and-paste DNA-only transposon is excised from the donor chromosome, a double-strand break is created in the vacated chromosome. This break can be perfectly “healed” by a homologous end-joining reaction.
- Alternatively, the break can be rescaled by a nonhomologous end-joining reaction; in this case, the DNA sequence that flanked the transposon is often altered producing a mutation at the chromosome site from which the transposon was excised. Mutation also occurs at the target site.
Mechanism Of Retrotransposition: The basic mechanism of retrotransposition involves the following three steps:
- An RNA copy of the transposon is synthesized by the normal process of transcription.
- The RNA transcript is copied into DNA, which initially exists as an independent molecule outside of the genome. This conversion of RNA to DNA, the reverse of the normal transcription process, requires a special enzyme called reverse transcriptase. Often the reverse transcriptase is coded by a gene within the transposon and is translated from the RNA copy synthesized in step 1.
“How do transposons move within genomes? FAQ explained”
The DNA copy of the transposon integrates into the genome, possibly back into the same chromosome occupied by the original unit, or possibly into a different chromosome. The result is that there are two copies of the transposon, at different points in the genome.
Examples of Transposons: Transposable elements are widespread among both prokaryotes and eukaryotes. Thus, they have been studied in diverse organisms, including bacteria, fungi, nematodes, insects, plants, and mammals.
Bacterial Transposons
- Insertion sequence (IS). During the 1960s geneticists were puzzled by a set of unusual mutations which were occasionally found in operons of E.coli and other bacteria.
- These are called polar mutations because they affect not only the gene in which the mutation occurs but also other genes downstream of the mutation.
- Several types of polar mutations are known and often they introduce a transcription termination signal into the operon.
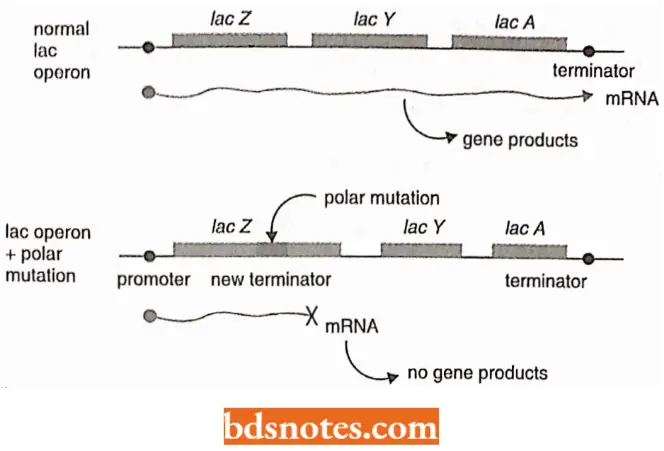
In contrast to other known mutations these polar mutations are always irreversible. Further, if a plasmid is introduced into a bacterium that carries a polar mutation of this type, then similar irreversible polar mutations suddenly appear in genes carried by the plasmid. It happens as though the mutation can jump from one DNA molecule to another.
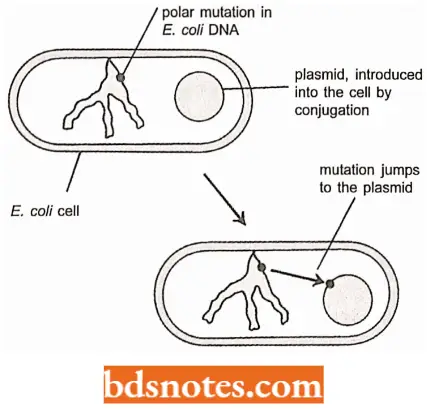
- The first hint of an explanation for this strange phenomenon appeared when the plasmids that had become mutated were studied. It was found that these plasmids had increased in size, generally a kilobase or more.
- When the DNA sequences of the mutated plasmids were examined it was discovered that the mutations were caused by the insertion into the target genes of new pieces of DNA, which were subsequently called insertion sequences or ISs.
- These insertion sequences possess the remarkable capability of being able to move around the genome from site to site by a process dependent on recombination and transposition. Moreover, when an IS transposases.
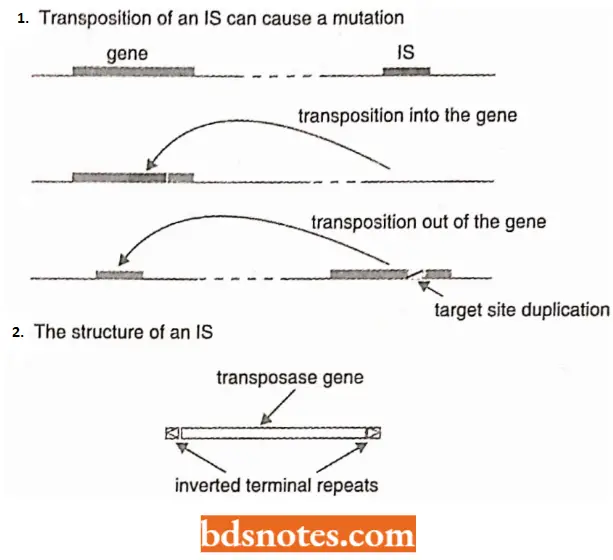
- IS elements are the simplest forms of transposons. They are so called because they were discovered as spontaneous insertions within bacterial operons that prevented transcription or translation of the concerned gene into which insertion has occurred.
- To a new site, it leaves behind a short duplication of the target site, usually between 5 to 15bp, which means that the gene that carried the IS probably remains mutated even after the IS element has departed.
- Various types of IS elements are found in E. coli. Each bacterium may contain several different IS types, each with a copy number of up to 20. Transposition does not occur so frequently, and a single cell may undergo a thousand cell divisions before one of its insertion sequences transposases.
- Insertion sequences can transfer between bacteria during conjugation, and can also transfer between bacteria during conjugation and can also transfer between related species.
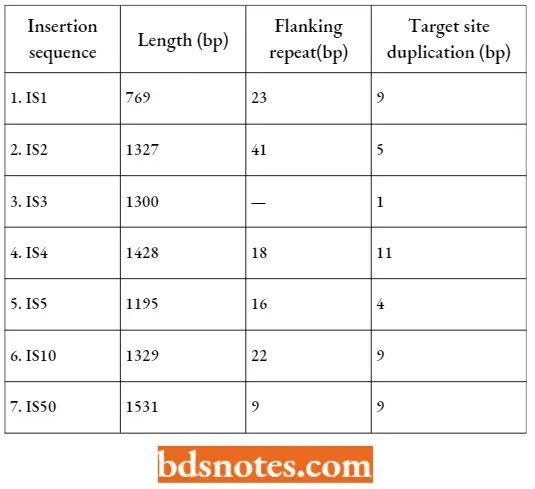
- Most of each IS element is taken up by a single gene (except IS1 which has two) coding for a transposase enzyme that catalyzes the transposition event. The IS element has at its ends a pair of inverted repeats between 9 and 41 bp in length, depending on the IS type.
Insertion Sequences Of E. coli:
“Early warning signs of gaps in understanding mobile genetic elements: Common questions”
Inverted Repeats:
Consider the following sequence of a DNA molecule:
5’— AG G CAT — 3′
3′ — TCCGTA — 5′
An inverted repeat of the above DNA sequence will be:
5’— AGGCATATGCCT — 3′
3’— TCCGTATACGGA — 5′
Repeats may occur adjacent to one other, as shown here, or may flank a different DNA sequence, as with the repeats at the ends of insertion sequences and Tn-3 type transposons.
- Composite transposons. They consist of two insertion sequences flanking an internal region which usually includes at least one gene, often coding for resistance to an antibiotic.
- Transposition occurs because of the transposase enzyme coded by IS elements. There are several different composite transposons, some over l0 kb in size. Often these types of elements are carried by the plasmids.
Composite And Tn3 – Type Transposons Of E. coli:
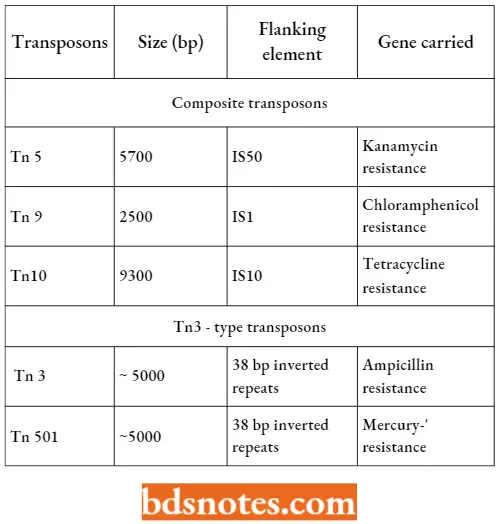
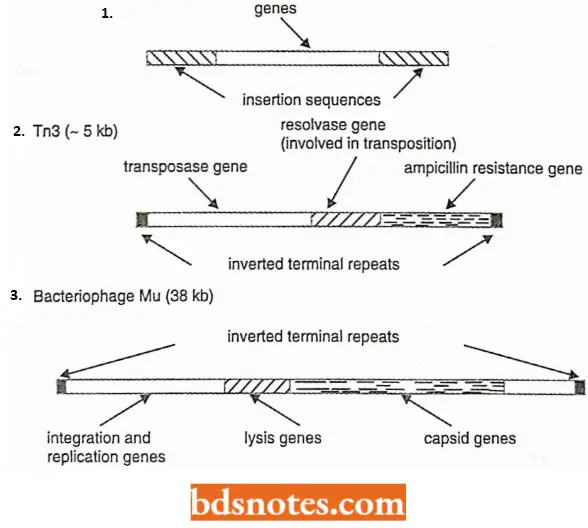
- Tn3 – type transposons. They are not flanked by IS elements but are still able to transpose because they carry a different transposase enzyme.
- Transposable phages. Certain phages such as Mn and D108 transpose as part of their normal infection cycle.
- This means that besides inserting themselves into the host DNA (like phage λ), the genomes of these phages can move about from place to place in the host DNA (unlike phase λ )
- Transposable phages may be transposons that have evolved genes for protective proteins, enabling them to exist outside the cell, or they may be ordinary phages that have learned to transpose (see Brown, 1998).
- Significance of bacterial transposable element. Transposons have no apparent function. Many evolutionary geneticists suggested that transposons are an evolutionary accident that once created, are self-maintaining.
- Since they may exist without a noticeable benefit to the host’s phenotype, transposons have been referred to as selfish DNA, i.e., they exist only to further their existence (propagation), contributing nothing to the cell as a whole.
However, it has been established that bacteria that carry a large number of transposons have a selective advantage over those with few transposons, possibly because transposons increase the mutation rate of the bacterium to adapt to changing environmental conditions.
- In higher organisms, the retroviruses, which include some viruses implicated in cancer as
well as the AIDS virus, resemble transposable elements. - In the laboratory, transposable elements are used as mutagens because they inactivate genes that they transpose into, or possibly modulate the expression of a gene downstream from an insertion site (for example., in lac operon of E. coli) in both cases, a phenotypic change indistinguishable from a mutation.
- Eukaryotic Transposons and Retroelements Eukaryotes contain a variety of a. Retrovirus transposable elements that were. originally discovered in maize and described as controlling elements.
- Insertion of a controlling element in a chromosome may affect the expression of adjacent genes. These elements may also produce deletions, duplications, inversions, and translocations at the sites in which they are present.
- Chromosome breakage is the common consequence of the presence of some transposable elements. In maize, the activities of controlling elements are regulated during development; they transpose and promote genetic rearrangements at characteristic times and frequencies during plant development.
Eukaryotic transposable elements are of the following types:
- Transposons and
- Retroelements Transposons are comparable to bacterial transposons, and have no life outside the genome, for example., controlling elements of maize.
RNA transposons or retroelements are features of eukaryotic genomes but have not so far been discovered in prokaryotes. They have attracted great attention because of their close similarity with free-living viruses, called retroviruses.
“Asymptomatic vs symptomatic effects of ignoring plasmid and transposon principles: Q&A”
- These viruses include both benign forms and also virulent types such as the human immunodeficiency viruses (HIV) that cause AIDS. These structural relationships are illustrated in and are of the following types:
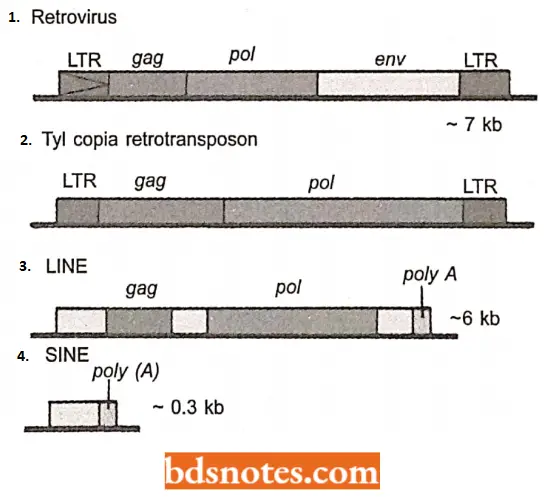
- Retroviruses. These are exogenous viruses whose genomes are made of RNA. Their genetic organization has been shown in. They infect many types of vertebrates.
- Once inside a cell, the RNA genome is copied into DNA by the reverse transcriptase enzyme specified by the viral pol gene, and the DNA copy integrates into the host genome.
- New viruses can be produced by copying the integrated DNA into RNA and packaging this into virus coat proteins, the latter coded by the env genes on the virus genome.
- Endogenous retroviruses (ERVs). These are retroviral genomes integrated into vertebrate chromosomes. Some are still active and might, at some stage in a cell’s lifetime, direct the production of exogenous viruses, but most are decayed relics that have lost the capacity to form viruses (Patience et al, 1997).
- These inactive sequences are interspersed repeats of the genome and they are not capable of additional proliferation.
- Retrotransposons. These elements have sequences similar to ERVs but are features of non-severbrate eukaryotic genomes (i.e., plants, fungi, invertebrate and microbial eukaryotes) rather than vertebrates.
Retrotransposons are of many types and each has very high copy numbers in some genomes. Most of the interspersed repeats in maize are retrotransposons; in maize, these elements form almost half of the genome. There are two types of retrotransposons :
- Ty3 / gypsy family. The members of this family have the same set of genes as an ERV, for example., the T3 genes of yeast, and the gypsy gene of Drosophila.
- Ty1 copia family. The members of this family lack the env gene. Both of these types of retrotransposons can transpose but the absence of the env gene means that the Tyl/copia group cannot form infectious virus particles.
- In fact, despite the presence of env in the Ty3/gypsy genome, some of these elements can form viruses and hence should be regarded as invertebrate retroviruses.
- Although technically they are interspersed elements, retrotransposons are sometimes found in clusters in a genome sequence as a result of the presence of preferred integration sites for transposing elements.
- The three types of retroelements described so far are LTR elements, as they have long terminal repeats at either end which play a role in the transposition process. Other retroelements do not have LTRs.
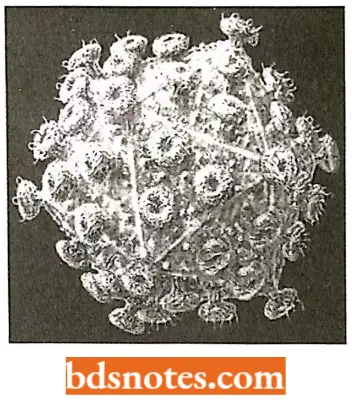
These are called retroposons and in mammals include the following:
- LINEs (Long Interspersed Nuclear Elements) They contain a reverse-transcriptase-like gene probably involved in the retrotransposition process (Fig 4.14C). An example is the human mobile element LINE – 1 which is 6.1 km long and has a copy number of 516000 in the human genome.
- SINEs (Short Interspersed Nuclear Elements) They do not have a reversed transcriptase gene but can still transpose, probably by “borrowing” reverse transcriptase enzymes that have been synthesized by other retroelements.
The commonest SINE in the human genome is Alu, which has a copy number of over one million. All seems to be derived from the gene for the 7SL RNA, a non-coding ScRNA (= 1 small cytoplasmic RNA) involved in the transport of proteins around the cell.
- The first Alu element may have arisen by the accidental reverse transcription of a 7SL RNA molecule and integration of the DNA copy into the human genome.
- Some Alu elements are copied into RNA, providing the opportunity for the proliferation of the element.
Examples of Eukaryotic Transposons
Transposons in Maize: In maize, at least 14 different transposable systems are known, for example., En, As-Ds, mu, etc. Each of these elements is present in multiple copies in the maize genome.
These have the following general characteristics: Autonomous and nonautonomous controlling elements.
The controlling elements of maize may be grouped into two classes:
- Autonomous controlling elements; and
- Nonautonomous controlling elements.
Autonomous controlling elements can excise and transpose themselves and corresponding nonautonomous elements. Nonautonomous controlling elements cannot transpose; they transpose only when an autonomous member of the same family is present elsewhere in the genome.
- The nonautonomous controlling elements may be derived from autonomous controlling elements by the loss of mans-acting functions needed for transposition. The autonomous controlling elements of maize are Ac (‘activator’: V9 copies/genome), Mul (mutator; > 10 copies/genome), Spm (‘suppressor mutator’, and Dd (‘dotted’).
- The nonautonomous controlling elements are Ds (dissociator, released to Ac), dspm (defective Spm; related to Spm), and(‘inhibitor’; related to En). The nonautonomous controlling elements possess the sites such as terminii, on which transpoases act.
- They also have defective ax-acting functions which make the elements permanently stable (since they can no longer be activated to transpose).
- Change of phase. Autonomously controlling elements may be reversibly inactivated during certain phases of plant development; this is called a ‘change of phase’. The phenomenon of change of phase is heritable but relatively unstable so that controlling element cycles between active and inactive states during development.
The inactivation of both,4c and Mil seems to be due to cytosine methylation in the target sequence CAG/GTC. Both Ac and Mu elements have several such target sites in each element. There may be self-regulative controls of transposition; an increase in the frequency of its transposition.
Examples of transposons of maize are the following: En controlling element. The En element (En = enhancer) codes for a single enzyme called transposase, which is believed to induce its transposition.
- Deletion of the En element during coding for this enzyme results in the element I (‘inhibitor’); / element is present in several copies in each maize line but it is non-transposable in the absence of En which acts as the signaling for.
- A similar situation exists for the Ac – Ds system maize chromosome, it may generate three-base duplication, for example., ATA, ATA, etc., in the gene from which it moves out; produce an inversion for a segment of the gene; or even cause duplication of the entire gene.
- Given these genetic effects, transposable elements are often referred to as genome reorganizers.
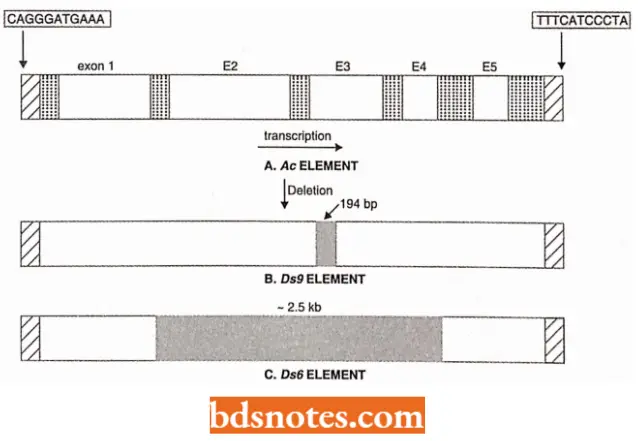
- Ac-Ds controlling elements. Ac (activator) controlling elements are 4,563 bp long, and encode a single mRNA of 807 codons (which encode transposase enzyme) from a gene having 5 exons.
- They have 11 bp inverted repeats at their ends. Ac elements generate repeats of 8bp at the sites of their insertion. The nonautonomous counterparts, of Ac elements occur in a variety of forms, for example., Dsl, Ds2dl, Ds2d2, Ds6, Ds9, etc., all of which contain the same 11 bp inverted repeats at their ends. Ds (= dissociation) elements are shorter than Ac, and the length of the deletion varies from merely 194 bp in Ds9 to over 2.5 kb in Ds6.
“Can targeted interventions improve outcomes using plasmid knowledge? FAQs provided”
Non-autonomous controlling elements (Ds elements) are derived from autonomous elements by deletions or other changes that inactivate their transposase enzyme coding regions, but the sites on which this enzyme acts remain intact.
- Members of the Ds1 family, however, are different their only resemblance to Ac is that they possess the same 11 bp terminal inverted repeats. Dsl controlling elements are short and were possibly not derived directly from Ac elements.
- Like the bacterial composite transposon such as Tn10, the transposition of Ds elements occurs by a nonreplicative mechanism. The recipient site is frequently on the same chromosome as the donor site and often quite close to it.
- In them, transposition almost always occurs after DNA replication, and usually, only the two copies produced by replication undergo transposition. The basis for this may be similar to that for TnlO and could involve asymmetrical methylation sites as C* GTG / G* CAC.
- After replication, the daughter DNA duplexes will have different methylated sequences, viz., C* GTG / GCAC and CGTG / G* CAC; one of these sequences may be recognized by the transposase enzyme more efficiently than the other.
- The donor site suffers a break, which could lead to a simple loss of the chromosome fragment distal to the centromere (hence lacking a centromere) or produce a breakage-fusion-bridge cycle.
- A loss of acentric fragment allows the expression of recessive alleles present on the corresponding region of the homologous chromosome. The breakage-fusion-bridge cycle, however, continues through the future generations causing changes (deletion, duplication) in the affected chromosome.
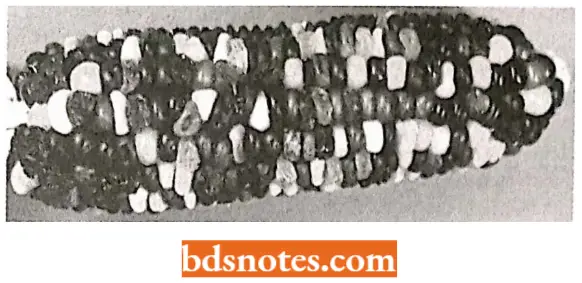
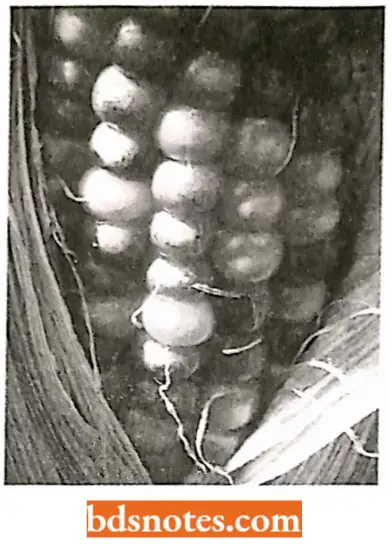
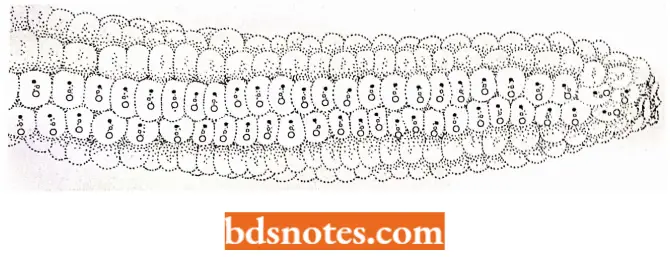
How purple spots are produced in maize kernels? Ds element affects the phenotype (in maize) by blocking expression of the genes it transposes into as well as causing JigD the loss of alleles in acentric chromosome fragments lost when Ds breaks its chromosome.
- In the colored figure, we see (a hive kind of kernels in the ear of corn purple, bronze without purple spots, and bronze with purple spots.
- The purple kernels result from the presence of dominant functioning alleles providing enzymes in the pathway for purple pigment.
- In those kernels that are bronze without spots, Ds elements have transposed into both copies of the B\ locus, disrupting the pigment pathway. Without the Ac element present, the Ds elements remain in place and the kernels are a uniform bronze color.
- In those bronze kernels with purple spots, the Ac element has entered the genome in the genetic cross. In the presence of Ac, Ds leaves its site in some of the cells, restoring the activity to the B2 locus.
- This restored activity results in purple spots in those cells and their progeny with the functioning B2 allele (Taniarin, 1996).
- Purple spots differ in size based on the time of excision of the Ds element during the development of the kernel: early yields large spots; late yields small spots.
How Ac-like Mobile Element Produces Mendel’s Wrinkled Peas? More recent work on transposable elements in plants has led us full circle to the union of Gregor Mendel’s observations with molecular genetics. Recall that, early in his work, Mendel investigated the inheritance of round and wrinkled peas.
- The two phenotypes are produced by alleles of a single gene, rugosus. It is now known that the wrinkled phenotype is associated with the absence of an enzyme, starch-branching enzyme (SBEI), that controls the formation of branch points in starch molecules.
- The lack of starch synthesis leads to the accumulation of sucrose and a higher water content and osmotic pressure in the developing seeds. As these seeds mature, those that are wrinkled (genotype rr) lose more water than do the smooth seeds (RR or Rr), producing the wrinkled phenotype.
- The structural gene for SBEI has been cloned and characterized in both wild-type and mutant genotypes. In the RR genotype, the SBEI protein is non-functional, presumably because the SBEI gene is interrupted by a 0.8-kb insertion, resulting in the production of an abnormal RNA transcript.
- The inserted DNA has 12-bp inverted repeats at each end that are highly homologous to the terminal sequences in the transposable element Ac from maize and to other Ac-like elements from snapdragons and parsley.
- Terminal repeated sequences and the genetic information encoding a transposase enzyme appear to be universal components of transposons in all organisms studied.
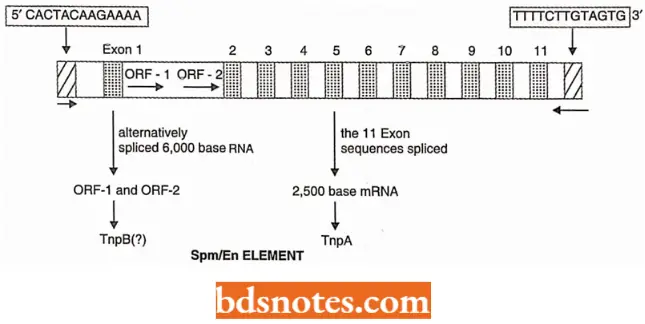
Spm-dSpm controlling elements: The Spin (suppressor mutator) elements have 13bp inverted repeats at their termini and have two genes. Gene tnpA has 11 exons that are spliced to produce a 2,500-base mRNA, which encodes the TnpA protein.
- Protein TnpA binds to a 12bp consensus region of Spin. The action of protein TnpA is required for excision, but the excision itself is affected by a hypothetical protein called TnpB.
- The protein TnpB is presumably coded by a 6000 base mRNA produced by alternative splicing of tnpA transcript; this mRNA contains the two open-reading frames, ORF – 1 and ORF – 2, both are present in the first large intron of gene tnpA.
- ORF – 1 and ORF – 2 are present in an alternatively spliced RNA of 6,000 bases; this RNA is present at ~1% of type mRNA. Protein TnB protein presumably binds the 13 bp terminal inverted repeats and cleaves the termini for transcription.
- A group of Spin derivatives are weakly autonomous; they are denoted as Spm-w. These controlling elements may show deletion in ORF – 1 and ORF – 2 regions; this indicates that the TnpB function during transposition may be bypassed or substituted.
- The dump controlling elements show deletion of tnpA exon sequences and the length of deleted sequences varies greatly.
Dotted (dt ) element. The dotted (Dt) element was discovered by Marcus Rhoades in 1938 when from a maize’s ear of pure breeding pigmented genotype of maize among kernels a dihybrid segregation ratio of 13 : 3: 1 (i.e., 13 pigmented : 3 dotted: 1 colorless) was obtained.
This was surprising because no segregation was expected. The genetic analysis suggested that the original phenotype (full color) was controlled by two genes with the genotype A1, A1 dt dt and that two mutations (A1 → a1 and dt → Dt) gave the genotype A1 a1 Dt dt which gave the following phenotypic and abbreviated genotypic ratio: 12/16 pigmented (A1Dt = 9/16; A1Dt = 3/16): 3/16 dotted (a1Dt): 1/16 colorless (a1 dt).

“Differential applications of simple vs composite transposons: Questions answered”
- The dolled phenotype (appears in kernels and leaves of maize) in a1Dt, would probably result from the reverse mutation ( a1→A1 ) in some of the somatic cells, and the frequency of this mutation should be unusually high.
- This led to the discovery of a1 as an unstable mutant allele (its instability is dependent on Dt) showing a high rate of reverse mutation.
- It was also shown that reverse mutations have occurred, they are stable, reverted At being transmitted to the progeny in stable form. It means that Dt causes mutations in a1 to A1, which remain stable.
- Trnusposons In Drosophila The genome of Drosophila melanogaster has several transposable elements, for example., P factor. P factor element.
- In D. melanogaster, F1 hybrids derived from certain strains show a series of defects, including a high mutation rate, high frequency of chromosomal aberrations, nondisjunctions, and sterility.
- The phenomenon by which such abnormal hybrids are produced is called hybrid dysgenesis. For example, when P (paternal contributing) type males are mated to M (maternal contributed) type females, the F1 progeny are sterile, while those from the reciprocal cross (P female x M male) are normally fertile.
- The P-type flies have a large number (30 to 50 copies) of P factors (the transposable elements) which become active in the cytoplasm of M flies. The active P factors transpose readily creating chromosome breaks and new mutations (for example., hybrid dysgenesis).
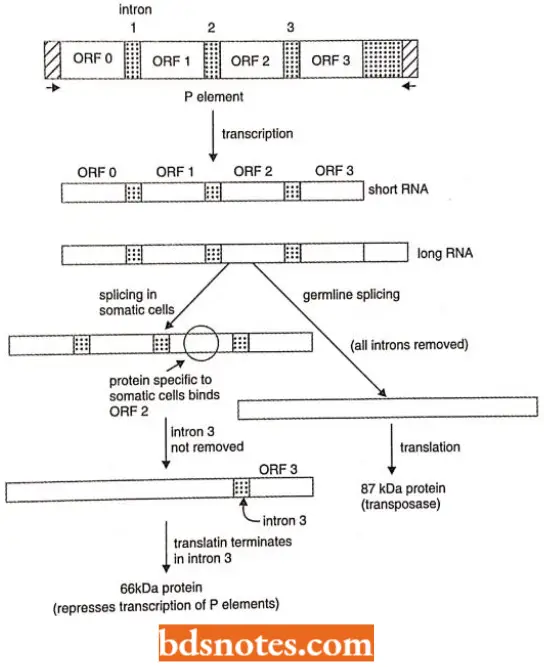
- The P elements vary in length, the longest being ~2.9 kb. They all have inverted terminal repeats of 31 bp and generate direct repeats of 8 bp in target DNA.
- The full-length P factors have 4 ORFs (= open reading frames), while their shorter types arise from internal deletions in them.
- The shorter P elements arise more frequently so about 2/3 of the elements present in P males are of this type.
- The transposition of P elements is nonreplicative like that of Tn10 (of E. coli), but the breaks in donor chromosomes are often repaired by using the corresponding sequence from homologous chromosomes.
- Transposition requires ~ 150 bp of terminal DNA and the transposase enzyme binds to lObp sequences that are adjacent to the 31 bp inverted terminal repeats.
- Further, the transposition of P elements is tissue-specific in that it occurs only in the germline cells; it begins at the stage when rapid cell division starts in the germline.
This tissue specificity occurs due to the tissue-specific alternative splicing of P elements transcripts as follows:
Germline splicing: It is a defective splicing pattern in which the 3 introns separating the 4 ORFs are spliced out generating an mRNA that encodes 87 kDa transposase enzyme. This enzyme induces P-element transposition.
Somatic splicing: It is induced by a protein, present in somatic cells of Drosophila. This protein binds to sequences in ORF-2 and prevents the removal of the 3rd intron, which has a termination codon.
The mRNA generated by this splicing produces a 66 kDa protein that does not contain information from ORF-3. This protein represses the transcription of P elements, thus, it stops transposition and forms the basis for reciprocal differences seen in P x M crosses of Drosophila.
“Steps to incorporate AI into analyzing plasmid and transposon functions: Questions and answers”
P cytotype and M cytotype: Drosophila of the P strain contains P elements in their genome (P chromosomes) and their cytoplasm has the 66 kDa protein; the cytoplasm of such flies is called P cytoplasm and they are said to have P cytotype.
- In contrast, flies belonging to the M strain do not have P elements in their chromosomes (M chromosomes) and lack the 66 kDa protein in their cytoplasm. These flies are said to have M cytoplasm that gives rise to the M cytotype.
- The 66 kDa protein is highly stable and is present in very large quantities in the eggs so that the P cytotype effect on the transposition of P elements may continue for more than one generation.
- Thus, when P males are mated to P females, or M males are mated to P females, the eggs contain the 66 kDa repressor protein.
- The transcription of P elements, therefore, remains suppressed and there is no transposition either in somatic or germline cells.
- But when M females are crossed to P males, the eggs lack the 66 kDa protein. P elements are, therefore, transcribed to produce the 66 kDa protein in somatic cells which suppresses further transcription of P elements.
- But in germline cells, defective splicing of P element transcripts leads to the production of transposase; this helps in P element transposition and hybrid dysgenesis. The P element is highly invasive when placed into a new population.
- The source of the P element seems to be another species of Drosophila. Since hybrid dysgenesis reduces inbreeding, it helps in specification by creating breeding barriers.
- Thus, multiple hybrid dysgenesis systems may develop in different populations; this would lead to the inability to intermate and therefore to specification.
Examples Of Retrgposons (Retroelements)
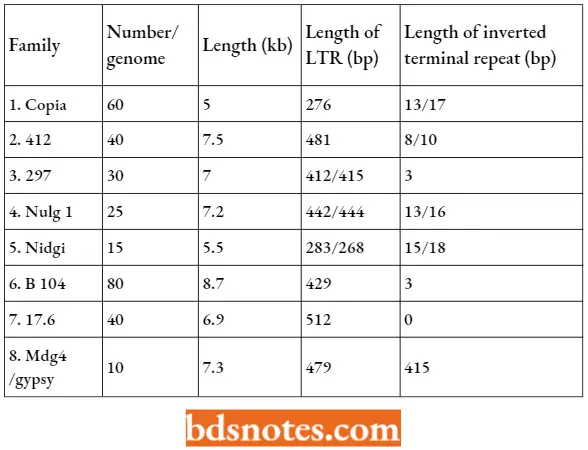
Copia elements in Drosophila. The Copia elements are 5000 bp long, have direct terminal repeats of 276 bp, they are always intact, sometimes they occur in circular form and they have a single long (4,227 bp) reading frame.
- Copia is present in 20 – 60 copies per genome depending on the strain of the fly and is represented by a large number of closely related sequences.
- These sequences code for about abundant poly (A)+ mRNAs. The name Copia is based on the presence of this sequence in copious amounts in the cellular RNA of Drosophila.
- In cultured cells, the number of copia elements per genome increases up to 2 to 3 times the number found in vivo.
- In cultured cells, the rate of transposition increases to 10-3 to 10-4 events per generation. A direct repeat of 5bp of target DNA is generated by the insertion of a Copia element.
- Each of the terminal direct repeats of Copia elements is identical, indicating their relationship with retroposons. The single Copia ORF has homologies with gag and pol sequences of retroviruses.
Properties Of Some Families Of Copia Like Elements:
- The mRNAs from Copia sequences are translated into a variety of proteins in vitro, ranging from 18 to 51 kDa, whose functions remain unknown.
- The Copia elements must have originated from retroviruses and they can transpose (but the mode of transposition is unknown).
- Integration of Copia element produces mutations, a classical example being white-apricot (wa) mutation for eye color caused by insertion of a copia-like element.
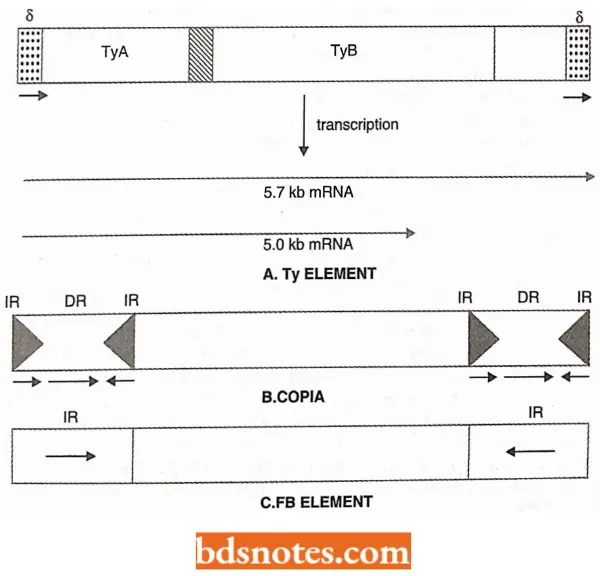
FB elements of Drosophila: The fold-back or FB elements range from 500 to 5,000 bp (base pair) and occur in about 30 copies per genome. They carry long (250 to 1,250 bp) inverted repeats at the termini.
- The repeats may be adjoining (FBI, FB5) or may be separated (FB3, FB4, FB7, FB8, FB9), but can always fold back to form stem and loop structures. FB elements cause mutations by insertion or their effect on gene expression (due to their presence near a control region).
- FB elements are also found flanking another class of transposon called TE (for example., TE1, TE28, TE77, TE98).
- TE1 was first identified due to the transposition of the white eye locus from the X chromosome to the second chromosome.
“Role of digital tools in improving precision with mobile genetic element tracking: FAQs explained”
- Ty dements in yeast. Yeast Ty (= transposon yeast) elements are an example of retroposons. Ty elements are very divergent. There are two main types of Ty elements, viz., Ty1 and Ty917. They transpose at a frequency of ~10-7 to 10-8 through an RNA intermediate.
- This RNA is reversed transcribed to yield a DNA duplex that becomes inserted into the yeast genome generating 5bp long direct repeats in yeast DNA. Such a transposition causes a mutation in the target gene due to insertion.
- Transpositions of T elements are controlled by genes within the T elements; they form noninfective virus-like particles, which accumulate within such cells in which transposition has been induced. Only some of the yeast T elements are active, while most of them have lost the ability to transpose.
- Each Tv element is 6.3 kb long and has 330 bp direct repeats at its ends (no inverted repeats or IRs are found in bacteria); these repeats are called delta (δ) repeats. All T elements share 8 repeats, a long region at the left end, another region in the center, and the short region adjacent to the right 8 repeats.
- In a typical yeast genome, there are ~30 copies of the Ty1 type and ~6 copies of the Ty 917 type element; in addition, there are ~100 copies of independent δ elements; these are called solo deltas. The sequences of δ elements vary greatly, but the two δ elements of a single TV element are identical.
- Thus, δ sequences seem to be similar to the LTR sequences of retroviruses. Although the two δ sequences of a T element are identical, a promoter is active in the δ repeats at the right end; this feature is also found in retroviruses.
The Ty elements can be excised by homologous recombination between the δ sequences, leaving behind also δ sequences in the host DNA. The Tv elements are transcribed into two species of poly + mRNAs of which one is 5 kb long and the other is 5.7 kb long.
- These mRNAs form > 75% of the total mRNA of a haploid yeast cell. Transcription for both mRNA species begins at ~95 bp from the left end of the Ty element. Further, the Ty elements have two open-reading frames (ORFs), which are read in different phases and overlapped by 13 codons. One ORF is TyA which encodes a DNA-binding protein.
- The other ORF is TB which contains regions that have homologies with reverse transcriptase, protease, and integrase sequences of retroviruses.
- The organization and functions of TyA and TyB are analogous to the behavior of retroviral gag and pol functions.
- Thus, TyB is expressed only when a frameshift event allows the termination codon at the end of TyA to be bypassed; as a result, TyB product is fused with that of Ty4. Moreover, the T elements comprise regions of homology within and between chromosomes.
- Recombination between Ty elements usually damages the chromosome either by deletion, inversion, or other more effective structural changes in chromosomes, for example., dicentric chromosomes.
- Recombination between Ty elements seems to occur in bursts and gene conversion occurs between Ty elements present at different locations.
- Quite interestingly, although both the δ repeats of an active Ty element have the same sequence, a promoter is active in one δ element, while a terminator is functional in the other. The Ty element behaves like a retrovirus that has lost its env gene so that it is unable to properly package its genome into virus particles.
“How do advancements in technology enhance plasmid and transposon research? Q&A guide”
Retrotransposons in plants: Retrotransposons are ubiquitous in plants. They make up to 50% to 80% of the genomic DNA (for example., in maize), thus, playing a major role in gene and genome evolution in plants.
- For example, LTR – retrotransposons are 100 bp to 13.5 kbp in size and possess the following coding sequences:
- Gag, encoding proteins involved in the maturation and packaging of RNA, and proteins necessary for the integration of retrotransposons into the genome pool, encoding reverse transcriptase (RT) and RNase H activities, needed for replication/ transposition to a new position on a chromosome.
- The Ty1-copia and Ty3-gypsy types of LTR- retrotransposons differ in the order of the above coding sequences, particularly the int and Rnase sequences. Among non-LTR retrotransposons, LINEs, though simpler and smaller in size (1000 bp to 6 kbp), still encode many of the proteins encoded by the LTR retrotransposons.
- For instance, they have gag and pol, but not int, and instead have endonuclease (en). SINEs do not encode any of these genes and are derived from RNA polymerase HI products such as tRNA.
- In a plant’s chromosome, the retrotransposons are either dispersed on all chromosomes or can be clustered in some regions such as telomeres, centromeres, pericentromeric regions, and also in the interstitial heterochromatic regions.
An individual retrotransposon can be present as a single copy or it may be represented by up to as many as 50,000 copies per genome. There can be dozens of different types of retrotransposons within a species (for example maize). Some of the known plant retrotransposons have been listed.
Some Selected Retrotransposons In Plants:
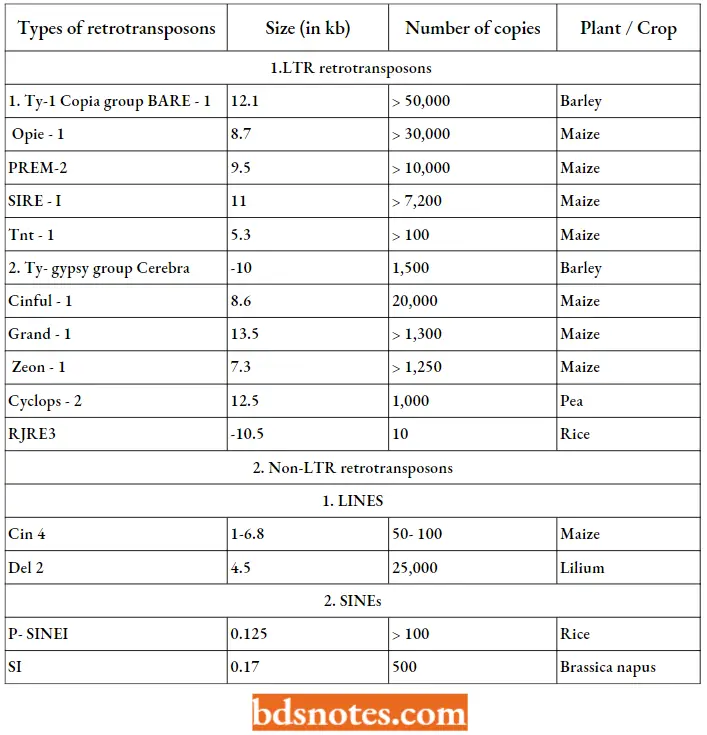
Tam 1 element in Snapdragon (Antirrhinum majus): In snapdragons, the Tam 1 element is responsible for the pattern of color seen on the petals of some variegated strains and is 17 kb long. Taml element is found in an unstable mutant, Nivea recurrent, where Taml is present in the region of the chalcone synthase gene and causes 3bp target site duplication.
- Tamil element also has 13bp inverted repeats at its termini, which are identical to those in Spml8 of maize. Other transposable elements of this series are numbered Tam2 to Tam10. Putative transposable elements are also suggested to be present in soya beans.
- Interspersed Repeats and Transposable Elements of Human Genome The draft sequences have shown that approximately 62% of the human genome comprises intergenic regions, the parts of the genome that lie between genes and which have no known function.
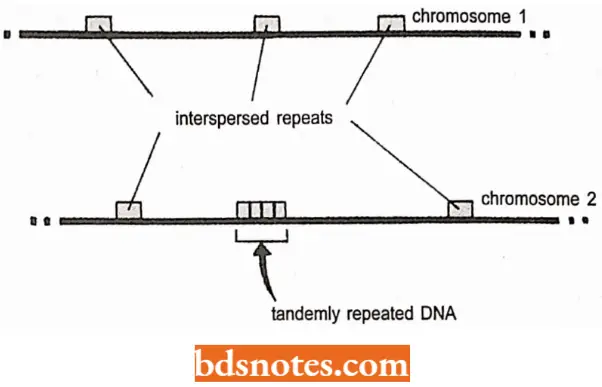
- These sequences used to be called junk DNA but the term is falling out of favor of modern molecular biologists. Recently, the bulk of the intergenic DNA is made up of repeated sequences of one type or another:
- Genome-wide or interspersed repeats, whose individual repeat units are randomly distributed around the genome; tandemly repeated DNA, whose repeat units are placed next to each other in an array.
- Interspersed repeats include four types of mobile elements: SINEs, LINEs, LTR elements, and DNA transposons. An interesting feature of these interspersed repeats is that each type appears to be derived from a transposable element, a mobile segment of DNA that can move around the genome from one place to another.
- Many of these mobile elements leave copies of themselves when they move, which explains how they propagate and become common throughout the genome.
- There are two main classes of transposable elements: those that transpose via an RNA intermediate and those that do not. LINEs, SINEs, and LTR elements are examples of the first class and DNA transposons are examples of the second class.
“Early warning signs of outdated methods in plasmid and transposon studies: Common questions”
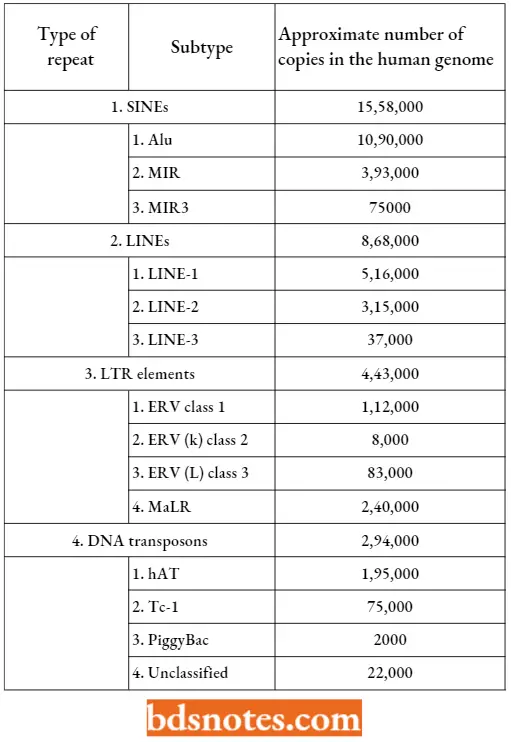
Just for example, SINEs form the most numerous genome-wide repeats of the human genome. They comprise three subtypes: Alu element (approximately 1,090,000 copies in the genome), MR (393,000 copies), and MR3 (75,000). Altogether genome-wide repeats make up 44% of the draft genome sequences, some 1400 Mb of DNA.
The Types Of Genome-Wide Repeats In The Human Genome:
Mammalian Retroposons: Mammals contain a variety of retroposons; most of these belong to the following two families :
- LINEs (Long Interspersed Elements): They are derived from transcripts of RNA polymerase 2.
- SINEs (Short Interspersed Elements): They are derived from transcripts of RNA polymerase 3.
LI retroposon is a good example of a LINE sequence. It is found in 20 to 50 thousand copies per genome. A typical LI element is ~6,500 bp long, terminating in an A-rich region.
- It appears that the LI element originated as a mobile gene coding for its transposition as its reading frame homologous to reverse transcriptase.
- The LINEs family shows great variation among individual members, but the members within a species are relatively homologous compared to those within species.
- In the 6,581 bp LI element, two open reading frames of 1,137 (ORF1) and 3,900 (ORF2) bp are present.; these reading frames overlap by 14 bp. ORF2 has a region homologous to the reverse transcriptase encoding sequence.
- Transcripts of this LI sequence are found in the cells. The LINE sequences originate from RNA polymerase 2 transcripts; as a result, they lack promoter sequences and are inactive.
- Since these sequences usually have the features of the mature transcript, they are called processed pseudogenes (these are different from pseudogenes).
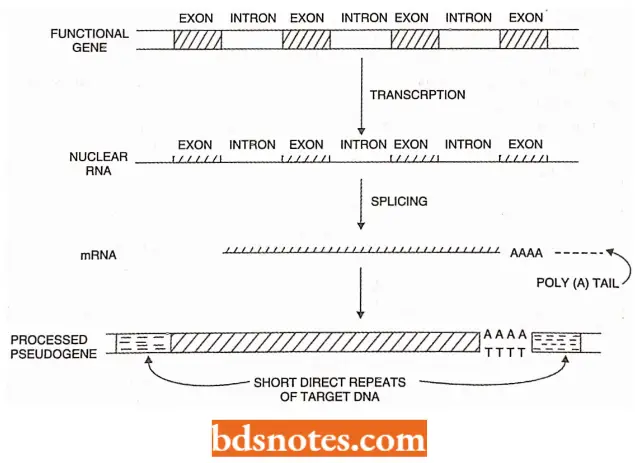
“Asymptomatic vs symptomatic effects of ignoring new trends in mobile genetic elements: Answered”
- This A-processed pseudogene is a pseudogene that results from integration into the genome
of a reverse-transcribed copy of mRNA pseudogenes have their 5′ ends having exact similarity with mRNA, many of them have exactly joined exon sequences and they usually end in a short 3′ stretch of AT base pairs. - This 3′ stretch is probably derived from the poly tail of the mRNA. Presumably, these pseudogenes have originated from a mobile gene encoding its transposition.
- It indicates that LI elements belong to the viral superfamily of retroposons. The processed pseudogenes are flanked on either side by a short direct repeat, each of which is presumed to have been generated by a transposon-like insertion event.
- The Ah1 family is a good example of the SINE sequence of the human genome. These sequences are so named because they all are cleaved at a single site located at 170bp by the restriction enzyme Alul.
- The sequence of the Alu family is ~300 bp long, occurs in ~300,000 copies per human genome, and is widely dispersed in her genome.
The human Alu family seems to have originated by a 130bp tandem duplication with an unrelated sequence of 31 bp inserted into the right half of the dimer. The Alu sequence is related to 7SL RNA, which is a component of the signal recognition particle.
- The 5′ terminal 90 bases of 7SL RNA are homologous to the left end of the left half (130 bp) of an Alu element, while the 40 bases at the 3′ -end of 7 SL RNA are homologous to the right end of the left half of Alu sequence; the central 160 bases of 7 SL RNA are absent from the Alu sequence.
- The genes for 7 SL RNA are transcribed by RNA polymerase III, and the SINEs are derived from transcripts of this RNA polymerase III. The Alu sequences likely originated from the transcripts of 7 SL RNA genes by a deletion of the central 160 bases of this RNA.
- The presence of multiple copies of the Alu sequence in a single RNA molecule can produce a secondary structure in the RNA. The presence of Alu sequences in the form of inverted repeats seems to be responsible for most of the secondary structures found in mammalian nuclear RNAs.
- Members of the Alu family resemble transposons in being flanked by short repeats of target DNA; the lengths of these DNA repeats are different for individual members of the family. Alu sequences have a 14 bp region that is almost identical to a sequence present at the origin of replication of approval viruses such as SV40 and hepatitis B virus.
Plasmids IS Elements Transposons Questions And Answers
Question 1. The initial discovery of IS units in bacteria involved their presence upstream (5′) to three genes controlling galactose metabolism. All three genes were affected despite not being contiguous with the IS unit. Explain as to why this occurs.
Answer: The IS probably occupied or interrupted the normal function of controlling regions related to the galactose genes.
Question 2. In maize, the recessive allele bz produces a lighter colour in the aleurone than does the dominant allele, Bz. Ears on a homozygous BJBz plant were fertilized by pollen from a homozygous BJBz plant. The resulting cobs contained kernels that were uniformly dark except for a few on which light spots occurred. Suggest an explanation.
Answer: Transposon-induced mutation or loss of a Bz allele in the endosperm.
Question 3. Sometimes solo copies of the LTR of a retrotransposon, such as gypsy, are found in Drosophila chromosomes. How might these solo LTRs originate?
Answer: Through crossing – over between the LTRs of a gypsy element.
Question 4. Escherichia coli cells that are resistant to kanamycin are infected with bacteriophage lambda. Phage particles that were purified from the lysed cells were used to infect a kanamycin-sensitive strain called Cl07 and cells that were lysogenised were picked from turbid plaques. These cells were subsequently plated on a medium containing kanamycin and several kanamycin-resistant colonies appeared. Suggest an explanation for the transfer of kanamycin resistance to the Cl07 strain.
Answer:
A transposon carrying the kanamycin resistance gene transposed from the original cells to some of the infected phages and subsequently conferred resistance on some of the lysogenised C107 cells, produced by the second infection.
“Can bioinformatics revolutionize plasmid and transposon classification? FAQs provided”
Plasmids IS Elements Transposons Multiple Choice Questions Answers
Question 1. Certain genetic elements that regularly “jump” to new locations, often inactivating the gene into which they become inserted, are called
- Episomes
- Heterochromatin
- Transposons
- Overlapping genes
Answer: 3. Transposons
Question 2. The jumping genes in maize were discovered by
- Beadle and Tatum
- Barbara McClintock
- H. G. Khorana
- T. H. Morgan
Answer: 2. Barbara McClintock
Question 3. Genes that confer antibiotic resistance on bacteria are located on
- Chromosomal DNA
- Plasmid
- RNA
- Polysome
Answer: 2. plasmid
Question 4. Plasmids found in cells of bacteria are molecules of
- DNA
- RNA
- Proteins
- None of these
Answer: 1. DNA
“Differential applications of traditional vs cutting-edge plasmid techniques: Questions answered”
Question 5. Which of the following is called the resistance transfer factor?
- Col – factor
- R – factor
- F – factor
- None of these
Answer: 2. R – factor
Question 6. Plasmid is a
- Double-stranded circular DNA
- Single-stranded DNA
- Extrachromosomal linear DNA
- None of the above
Answer: 1. Double-stranded circular DNA
Leave a Reply